Hauptinhalt
2) Aufklärung der Funktion von LDs während der HCV Replikation und ihre Rolle in anderen infektiösen Erkrankungen
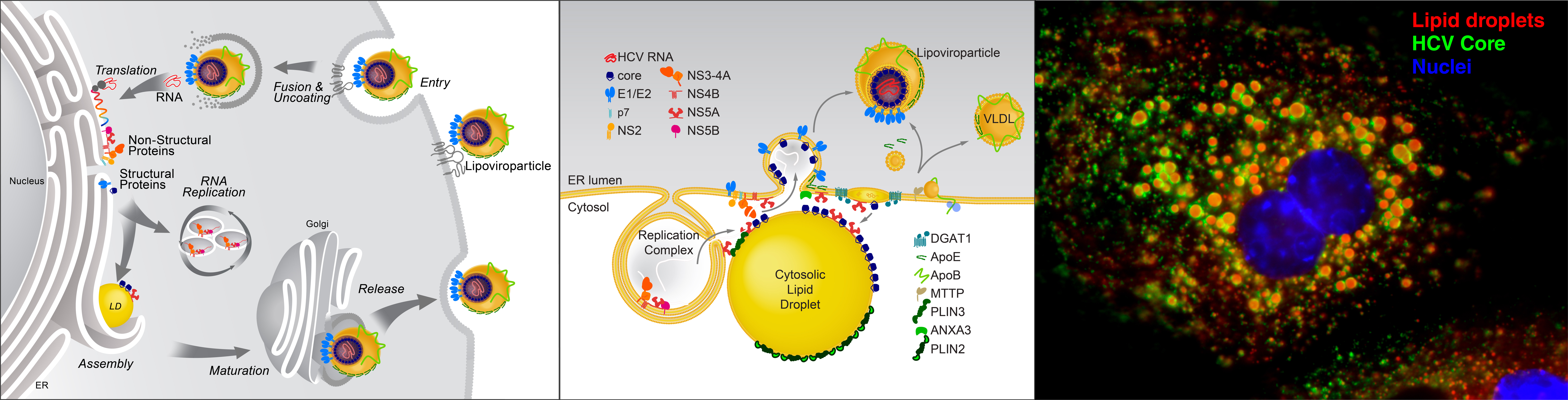
Schema der HCV Replikation und Morphogenese an lipid droplets (Modifiziert nach Herker and Ott, J. Biol. Chem., 2012). Das HCV Kapsidprotein Core lokalisiert in primären Hepatozyten zu lipid droplets.
Infektiöse HCV-Partikel bestehen aus einem umhüllten Kapsid, welches das positiv orientierte einzelsträngige RNA Genom enthält. Aufgrund der Assoziation mit Lipoproteinen und neutralen Lipiden werden HCV-Partikel auch als Lipoviropartikel bezeichnet. HCV infiziert fast ausschließlich menschliche Hepatozyten und repliziert im Cytoplasma der Wirtszelle. Das virale Genom kodiert für ein einzelnes Vorläufer-Polyprotein, das in die zehn viralen Proteine prozessiert wird: Die drei viralen Strukturproteine (das Kapsidprotein Core und die Hüllproteine E1 und E2), das Viroporin p7 und sechs Nichtstrukturproteine (NS: NS2, NS3, NS4A, NS4B, NS5A und NS5B). Die virale RNA Replikation findet in virusinduzierten vesikulären Strukturen statt, die einen hohen Anteil an Cholesterol und Sphingolipiden enthalten und Inhibitoren der Cholesterol- und der Sphingomyelinbiosynthese hemmen gleichzeitig die virale RNA Replikation. Diese Strukturen bestehen aus Einfach-, Doppel- und Mehrfachmembranvesikeln (membranous web) und es wird angenommen, dass sie die Replikationszentren vor der Erkennung durch pattern recognition Rezeptoren zu schützen. Zusätzlich finden sich in der Nähe dieser Membranstrukturen cytosolische LDs, die mutmaßlich als Assemblierungsplattform neuer HCV-Partikel dienen (Herker and Ott, Trends Endocrinol. Metab., 2011, Herker and Ott, J. Biol. Chem., 2012).
In unseren Arbeiten konnten wir zeigen, dass die LD-Biogenese notwendig für eine effiziente Virusproduktion ist. Diese Ergebnisse unterstreichen die essentielle Rolle von LDs während der viralen Replikation (Herker et al., Nat. Med., 2010). Mechanistisch erfordert die Translokation von Core und NS5A zu LDs die de novo Triglycerid Biosynthese, da Inhibitoren der Diacylglycerolacyltransferase-1 (DGAT1) den Transport der viralen Proteine zu LDs und die anschließende HCV-Assemblierung hemmen (Herker et al., Nat. Med., 2010, Camus et al., J. Biol. Chem., 2013). Zusätzlich verringert das Kapsidprotein Core an der Oberfläche von LDs ihren Abbau und induziert so sowohl in Zellkultursystemen als auch in Lebern von Mäusen die Ausbildung einer Steatose (Harris et al., J. Biol. Chem., 2011, Camus et al., J. Biol. Chem., 2014).
Mittels quantitativer LD-Proteomanalysen konnten wir zeigen, dass HCV die LDs von ihrer normalen metabolischen Funktion und Regulation abkoppelt. Außerdem haben wir das Wirtsprotein Annexin A3 (ANXA3) als neuen Wirtsfaktor für die Reifung von HCV-Partikeln identifiziert (Rosch et al., Cell Rep, 2016). Perilipin 2 (PLIN2/ADRP) ist das häufigste Protein an der Oberfläche von LDs in Hepatozyten und kürzlich haben wir die Folgen eines PLIN2-knockdowns im Zellkultursystem untersucht. Unsere Daten zeigen, dass LDs in PLIN2-defizienten Zellen in Doppelmembransäcken eingeschlossen sind und dadurch die infektiöse HCV-Partikelbildung nicht mehr unterstützen können (Lassen et al., J. Cell Sci., 2019).