Molecular Physiology of Plant-Fungal-Interactions (Voll Lab)
Plant pathogens fuel their metabolism by diverting organic and inorganic solutes from colonized host tissue, which is crucial for the successful establishment of the microbes in planta. Our group investigates metabolic processes in the interaction of plants with fungal pathogens and how the allocation of organic carbon and nitrogen sources between host and pathogen is controlled on the local and systems level. More specifically, we address the following questions:
1) How do the pathogens sense and respond to nutrient availability in the colonized host tissue? 2) Which host genes play a role for the redirection of nutrients to the pathogen? 3) How is the crosstalk between nutrient and defense signalling achieved in the host?
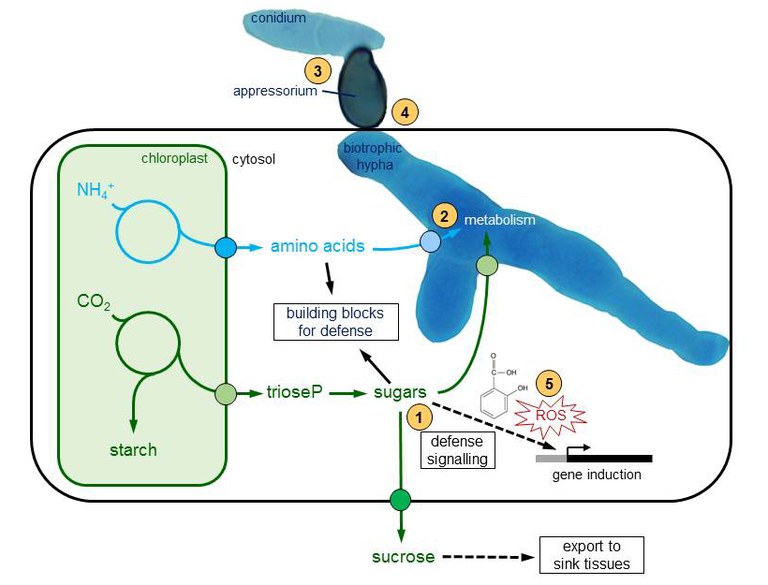
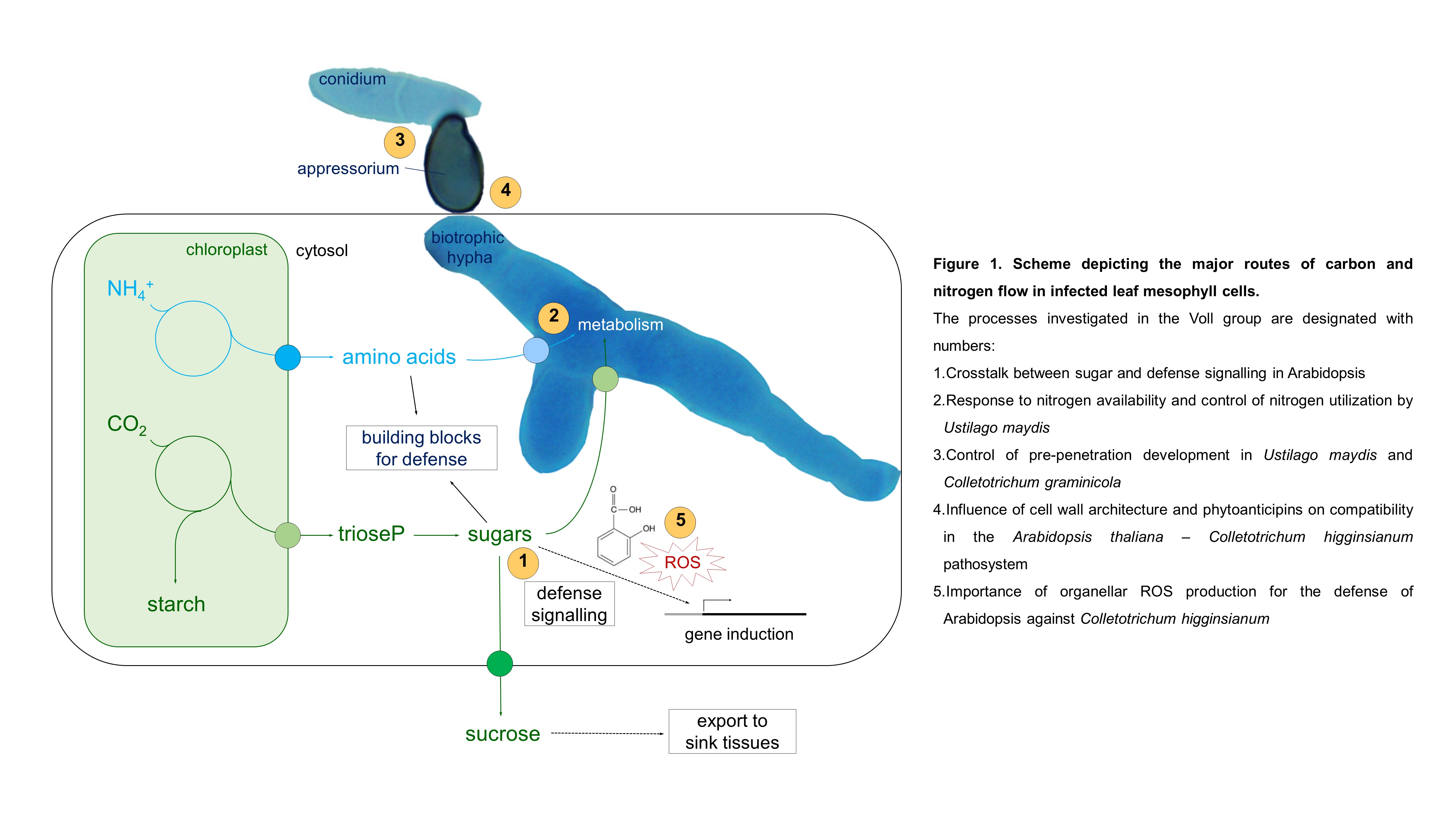
Biotrophic pathogens rely on an active provision of assimilated carbon (sugars) and nitrogen (amino acids) by living host cells (Figure 1). For their successful establishment, biotrophs need to suppress host defense immediately after penetration into the host organ and reprogram host metabolism to install high rates of assimilate flux into the fungal cytoplasm. Leaf pathogens generate strong local sinks for sugars and amino acids at the site of assimilate production, i.e. in photosynthetically active tissue.
Our group investigates metabolic reprogramming processes in host and pathogen during the interaction of plants with (hemi)biotrophic fungal pathogens (Figure 2). One central question guiding our work is how the allocation of organic carbon and nitrogen resources between host and pathogen (see Figure 1) is controlled on the local and systems level. To address specific questions, we have three pathosystems under study, in which the fungal partners are genetically tractable (Figure 2; for episodes from the individual fungal life cycles, please see Figures 3-5):
Scheme depicting the major routes of carbon and nitrogen flow in infected leaf mesophyll cells.
The processes investigated in the Voll group are designated with numbers:
1. Crosstalk between sugar and defense signalling in Arabidopsis
2. Response to nitrogen availability and control of nitrogen utilization by Ustilago maydis
3. Control of pre-penetration development in Ustilago maydis and Colletotrichum graminicola
4. Influence of cell wall architecture and phytoanticipins on compatibility in the Arabidopsis thaliana – Colletotrichum higginsianum pathosystem
5. Importance of organellar ROS production for the defense of Arabidopsis against Colletotrichum higginsianum
Host metabolism is highly dynamic during the interaction of plants with fungal parasites and most of these dynamic metabolic processes are highly relevant for the allocation of organic carbon and nitrogen building blocks. The flowing themes are currently investigated in the Voll group (see numbers in Figure 1 as a guide).
(1) We have found indication for a crosstalk between sugar and defense signaling in the interaction of Arabidopsis with Colletotrichum higginsianum. While reduced availability of carbohydrates interferes with the salicylic acid-dependent defense response of the host, ectopic carbohydrate accumulation in leaves activates salicylic acid signaling prematurely. It is yet unclear, how these sugar signals are generated on the spatiotemporal level, i..e. which sugar species are monitored in which cellular compartments to trigger a signal. We investigate these connections on the functional level, using reporter plants (see e.g. Figure 6) and mutants with altered sugar allocation, like the starchless p-pgm and adg1-1 mutants, or sweet11/sweet12 double mutants that exhibit hampered sucrose export from leaves.
(2/3) As a global regulator of Nitrogen Catabolite Repression (NCR), the Zn-finger transcription factor Nit2 controls the utilization of nitrogen sources in saprophytic conidia of the corn smut fungus Ustilago maydis. In the presence of favorable nitrogen sources like ammonium or glutamine, Nit2 represses genes required to utilize other nitrogen sources that are commonly less abundant or require more metabolic energy for their metabolization, e.g. nitrate, nucleobases or minor amino acids. Nit2 is also expressed at several growth stages of Ustilago maydis during maize colonization. U. maydis Nit2 mutants show a delayed induction of phytopathogenic growth on the surface of maize leaves (the infection stage depicted in Figure 4), and molecular data suggest that Nit2 is involved in the network regulating the dimorphic switch from saprophytic growth of haploid sporidia to phytopathogenic filamentous growth of dikarya that are formed after fusion of compatible sporida. Furthermore, we are interested in how Nit2 target genes are regulated during later phytopathogenic growth of Ustilago maydis in planta, when the host leaf gets massively colonized by intercellularly and intracellularly growing fungal hyphae. To this end, we investigate how U. maydis gene expression responds to nitrogen availability and quality during maize infection, and how Nit2 is involved in the adaptation to nitrogen availability. As a next step, we aim at identifying Nit2 target genes that are involved in nitrogen acquisition from the host and will resolve how they contribute to pathogenicity of U. maydis.
(3) We have screened for genes that weakly affect pathogenicity in the corn anthracnose fungus Colletotrichum graminicola. Among other genes, we identified an ubiquitin E2 ligase, CgUbc8, that is required for efficient glycogen storage during the development of fungal conidia. Consequently, penetration of maize leaves is hampered when mutant C. graminicola conidia that contain less glycogen reserves germinate to develop infection structures. We currently investigate the targets of CgUbc8 and how they interfere with glycogen production on the molecular and physiological level.
(4) Starchless Arabidospis mutants suffer from reduced nocturnal carbohydrate availability. This carbon limitation affects cell wall architecture in Arabidopsis, predominantly pectic polysaccharides. While the changes in cell wall composition caused by periodic carbon shortage in starchless mutants lead to reduced penetration resistance towards Colletotrichum higginsianum, other changes in pectin structure lead to improved penetration resistance. We try to discover the regulatory and physiological connection between the turnover of major storage carbohydrates, cell wall composition and resistance to fungal pathogens. We have also observed that penetration resistance towards non-adapted powdery-mildew fungi is impaired in barley plants with impaired starch biosynthesis, indicating that the underlying mechanism might be conserved between different plant systems.
(5) Reactive Oxygen Species (ROS) are important, widespread second messengers in eukaryotes. During the plant defense response, a burst of ROS, predominantly superoxide anions (O2•-) and hydrogen peroxide (H2O2), is produced in the apoplast upon recognition of invading microbial pathogens in order to defeat the intruder. In photosynthetically active cells, ROS are constantly produced in chloroplasts, peroxisomes and mitochondria, albeit at different rates. While the antioxidant system keeps the steady state level of ROS low, short term increases in organellar ROS production can serve as signals. Work in the Voll group has helped to refine the contribution of defense responses in response to chloroplastic ROS signals that are regulated by the transcription factor WRKY33 in Arabidopsis. Furthermore, recent results indicate that mitochondrial redox state rather than mitochondrial ROS production play a role for defense of Arabidopsis against Colletotrichum higginsianum.
Key methods used in our group are:
Metabolite quantitation of primary and secondary metabolites by microplate reader assays, Ultra-High Pressure Liquid Chromatography (UHPLC) and Ion exchange Chromatography
(IC)Analysis of host and fungal cell wall composition
Confocal Laser Scanning Microscopy (CLSM) of fluorescent reporter gene constructs in host and parasite (Figure 6)
Stable isotope labelling with 13C and 15N and flux analysis by LC-MS
Chlorophyll fluorescence imaging, gas exchange measurements and physiological bioassays (Figure 7)